| § | библиотека – мастерская – | Помощь Контакты | Вход — |
Данилова Н.Н., Крылова А.Л. Физиология высшей нервной деятельности. -- М.: Учебная Литература, 1997
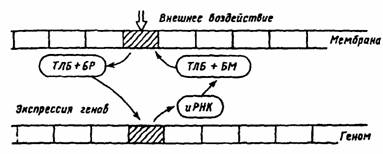
Стр. 157 Относительно механизмов модификации участков генома как феномена, ответственного за синтез и хранение код а «бе л ков обучения », Е.Н. Соколов [38; 39] предлагает модель. Согласно его гипотетической модели вся мембрана нейрона функционально проецируется на геном (рис. 26). Каждому локусу мембраны, содержащему специфический рецепторный белок, на геноме соответствует определенный участок экспрессированных или депрессированных генов. Воздействие медиатора или его спутника будет избирательно усиливать реактивность соответствующих ему рецепторных белков мембраны. Таким образом, каждый активированный участок мембраны выступает в форме «меченого белка». Белок-«метка» обладает специфической транслокационной группой и
Рш: 26. Гипотетическая схема отражения мембраны на геноме нейрона (по Т.Н. Греченко, Е.Н.Соколову, 1987). ТЛБ — транслокационный белок, БР — белок-регулятор, БМ — белок-модулятор, иРНК — информационная рибонуклеиновая кислота транспортируется к определенным участкам генома, вызывая избирательную экспрессию генов. В результате экспрессии определенных генов и при участии информационных молекул РНК происходит их транскрипция. Синтезируемый энзим (белок-модулятор) транспортируется белком к тому участку мембраны клетки, который был закодирован «маркерным» белком. Эта схема предполагает, что экспрессия генов, вызванная однократно, затем длительно удерживается, обеспечивая поддержание уровня синтеза белков, регулирующих работу рецепторов, задействованных нейромедиатором активного синапса (см. рис. 26). По этой схеме переход от привыкания к выработке следа памяти происходит по преи постсинаптическому механизму (см. рис. 24). В случае выработки нового навыка, условного рефлекса должна начать действовать другая комбинация синапсов и нейромедиаторов. Действует другой белок-модулятор, который переводит соответствующий рецепторный белок в реактивное состояние, и он может длительно присоединять к себе специфический медиатор (см. рис. 25). На пресинаптическом уровне кальций, активируя протеинкиназу, запирает калиевые каналы и тем самым усиливает вход кальция и выброс нейромедиатора в си-наптическую щель. На постсинаптическом уровне к активации натриевых каналов присоединяется вход кальция, включение протеинкиназы и выключение калиевых каналов, что повышает возбудимость нейрона. При участии генома нейрона усиливается синтез белков-модуляторов, влияющих неэффективность протеинкиназ, регулирующих закрытие калиевых каналов [10]. |
Реклама
|
||